Рибосомы - важные органеллы клетки, которые находятся на поверхности эндоплазматической сети. Строение рибосомы связано с синтезом белка.
Строение
Рибосома - это немембранная органелла, состоящая из двух частей - субъединиц. Рибосомы попадают на ЭПС или в цитоплазму из ядрышка через поры мембранной стенки ядра.
В зависимости от расположения рибосомы бывают двух видов:
- связанные - оседают на ЭПС;
- свободные - находятся в цитоплазме.
Субъединицы делятся на два типа - большие и малые. Каждая часть состоит из смеси нуклеиновых кислот и протеина, т.е. по химической структуре рибосома является нуклеопротеидом.

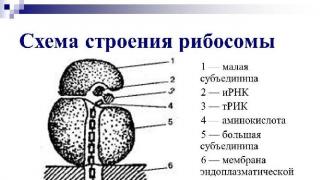
Рис. 1. Строение рибосом.
В состав рибосом эукариотической клетки входят четыре вида рибосомальной РНК (рРНК), различающихся количеством нуклеотидов:
- 18S - 1900 нуклеотидов;
- 5S - 120 нуклеотидов;
- 5,8S - 160 нуклеотидов;
- 28S - 4800 нуклеотидов.
18S-рРНК и 30-35 белков составляют малую субъединицу, остальные рибонуклеиновые кислоты и 45-50 белков входят в состав большой субъединицы. Большая субъединица прокариот включает два вида РНК, а малая - один.
В ядрышке субъединицы синтезируются по отдельности. Они собираются в месте в единую рибосому только для работы - синтеза белка, который происходит на матричной РНК. Субъединицы обхватывают мРНК, собираясь в комплексы, которые называются полисомами или полирибосомами.
ТОП-4 статьи которые читают вместе с этой

Рис. 2. Полисомы и мРНК.
По строению рибосомы животной клетки ничем не отличаются от растительной клетки. Однако клетки растений содержат значительно меньше рибосом, т.к. основную роль в обмене веществ играют хлоропласты.
Функции
Главная функция органоида - синтез белка.
Биосинтез белка включает несколько компонентов:
- мРНК;
- рРНК;
- полипептид;
- 20 аминокислот;
- ГТФ (гуанозинтрифосфат) в качестве источника энергии;
- рибосомальные белки;
- белковые факторы, регулирующие процесс.
Биосинтез происходит в два этапа:
- транскрипция - считывание и копирование информации с ДНК, образование мРНК;
- трансляция - синтез белка на рибосомах с помощью транспортной РНК (тРНК).
Матричная РНК - слепок, шаблон с ДНК, по которому рибосома синтезирует белок. Самая короткая рибонуклеиновая кислота - транспортная РНК - переносит аминокислоты к месту синтеза белка, выстраивая полипептидную цепь. При этом для каждой аминокислоты существует своя тРНК.
Процесс трансляции включает три фазы:
- инициацию - рибосома прикрепляется к началу мРНК;
- элонгацию - собственно синтез белка, образование полипептидной цепи;
- терминацию - высвобождение синтезированной цепи от рибосомы.
Элонгация происходит довольно быстро. За секунду полипептидная цепь увеличивается примерно на 20 аминокислот. Высвобождению цепи способствуют стоп-кодоны (УАА, УАГ, УГА) на мРНК. Данные кодоны не кодируют аминокислоты, и синтез на них заканчивается.
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ДОКЛАД
по дисциплине:
«Введение в естествознание»
«Рибосомы. Роль РНК в архитектуре
и функциях рибосом.»
Факультет: ФЕН
Группа: 7403
Студент: Романова Т. Е.
Новосибирск, 2007 г.
1. История исследований рибосомы
2. Строение и функции, разновидности рибосом
3. Принципы функционирования, роль РНК
4. Список
литературы
1.
История исследования рибосомы
Рибосомы впервые были описаны как уплотненные частицы, или гранулы, клеточным биологом румынского происхождения Джорджем Паладе в середине 1950-х годов. В 1974 г. Паладе, Клод и Кристиан Де Дюв получили Нобелевскую премию по физиологии и медицине «за открытия, касающиеся структурной и функциональной организации клетки». Термин "рибосома" был предложен Ричардом Робертсом в 1958 вместо "рибонуклеобелковая частица микросомальной фракции". Биохимические и мутационные исследования рибосомы начиная с 1960-х позволили описать многие функциональные и структурные особенности рибосомы. В начале 2000-х появились атомные структуры отдельных субъединиц, а также полной рибосомы, связанной с различными субстратами, которые позволили понять механизм декодинга (распознавания антикодона тРНК, комплементарного кодону мРНК) и детали взаимодействий между рибосомой, антибиотиками, тРНК и мРНК.
Рибосома - крупный внутриклеточный
макромолекулярный ансамбль, ответственный за синтез полипептидной цепи из
аминокислот (трансляцию); состоит из молекул РНК (т. наз. рибосомные
рибонуклеиновые кислоты, или рРНК) и белков.
Основная масса рибосом локализована в цитоплазме. В бактериальной
клетке рибосомы составляют до 30% ее сухой массы: на одну бактериальную клетку
приходится примерно 104 рибосом. В эукариотических клетках (клетки всех
организмов, за исключением бактерий и синезеленых водорослей) относительное
содержание рибосом меньше, и их количество очень сильно варьирует в зависимости
от белок-синтезирующей активности соответствующей ткани или отдельной клетки.
В эукариотической клетке все
рибосомы цитоплазмы (как мембрано-связанные, так и свободные) образуются в
ядрышке; считается, что там они неактивны. Эукариотическая клетка имеет также
специальные рибосомы в митохондриях (у животных и растений) и хлоропластах (у
растений). Рибосомы этих органелл отличаются от цитоплазматических размерами и
некоторыми функциональными свойствами. Они образуются непосредственно в этих
органеллах.
Различают два основных типа
рибосом. Всем прокариотическим организмам (бактерии и синезеленые водоросли)
свойственны так называемые 70S рибосомы,
характеризующиеся коэффициентом (константой) седиментации около 70 единиц
Сведберга, или 70S (по коэф. седиментации различают и
рибосомы других типов, а также субчастицы и биополимеры, входящие в состав Р.).
Их молекулярная масса составляет 2,5 · 10 6 , линейные размеры 20-25
нм. По химическому составу это рибонуклеопротеиды; они состоят только из рРНК и
белка (соотношение этих компонентов 2:1). Рибосомная РНК в рибосоме
присутствует в основном в виде Mg-соли (по-видимому,
частично и в виде Са-соли); магния в рибосоме до 2% от сухой массы. Кроме того,
в различных количествах (до 2,5%) могут присутствовать также катионы
аминов-спермина H2N(CH2)3NH(CH2)4NH(CH2)3NH2,
спермидина H2N(CH2)3NH(CH2)4NH2 и др.
Поскольку коэффициенты
седиментации зависят не только от молекулярной массы, но и от формы частиц,
седиментационные коэффициенты при диссоциации неаддитивны: так, например,
бактериальные рибосомы с молекулярной массой ~3*106 Дальтон имеет коэффициент
седиментации 70S, обозначается как 70S и диссоциирует на субъединицы 50S и 30S:
70S 50S + 30S
Рибосомные субчастицы содержат по
одной молекуле рРНК большой длины, масса которой составляет ~1/2 - 2/3 массы
рибосомной субчастицы, таким образом, в случае бактериальных рибосом 70S
субчастица 50S содержит рРНК 23S (длина ~3000 нуклеотидов) и субчастица 30S
содержит рРНК 16S (длина ~1500 нуклеотидов); большая рибосомная субчастица
кроме «длинной» рРНК содержит также одну или две «коротких» рРНК (5S рРНК бактериальных
рибосомных субчастиц 50S или 5S и 5.8S рРНК больших рибосомных субчастиц
эукариот).
Цитоплазма клеток всех
эукариотических организмов содержит несколько более крупные 80S
рибосомы. Их молекулярная масса около 4·10 6 , линейные размеры 25-30
нм, содержание белка в них значительно больше, чем в прокариотической рибосоме
(соотношение РНК: белок ок. 1:1). Рибосомная РНК 80S
также связана в основном с Mg и Са и с небольшим
кол-вом полиаминов (спермин, спермидин и др.).
Хлоропласты и митохондрии эукариотических
клеток содержат рибосомы, отличные от типа 80S.
Рибосомы хлоропластов высших растений принадлежат к истинному 70S типу. Митохондриальные рибосомы более разнообразны; их
строение находится в зависимости от таксономической принадлежности организма (т.е.
от принадлежности к определенному виду, роду или семейству). Напр.,
митохондриальные рибосомы млекопитающих существенно мельче типичных 70S Р.; коэффициент седиментации этих рибосом составляет около
55S (т. наз. минирибосомы).
Рибосомы из самых разнообразных
организмов имеют сходное строение. Они состоят из двух разделяемых субчастиц,
или рибосомных субъединиц. При определенных условиях (например, при понижении
концентрации Mg 2 + в среде) рибосома
обратимо диссоциирует на две субчастицы с соотношением их молекулярных масс
около 2:1. Прокарйотическая 70S рибосома диссоциирует
на субъединицы с коэффициентом седиментации 50S
(молекулярная масса 1,5·10 6) и 30S
(молекулярная масса 0,85·10 6). Эукариотическая рибосома разделяется
на субчастицы 60S и 40S. Две рибосомные
субчастицы объединены в полную рибосому строго определенным образом,
предполагающим специфические контакты их поверхностей.
Как прокариотические, так и
эукариотические рибосомы содержат две различные высокомолекулярные рРНК (по
одной на каждую субчастицу) и одну относительно низкомолекулярную рРНК в
большой субчастице.
Рибосомные белки большинства
животных представлены в основном умеренно основными полипептидами, хотя имеется
несколько нейтральных и кислых белков. Молекулярные массы рибосомных белков
варьирует от 6 тыс. до 60 тыс. г/моль. В прокариотической рибосоме малая
субчастица (30S) содержит около 20, большая (50S) - около 30 различных белков; в эукариотической рибосоме 40S субчастица включает около 30 белков, а 60S-около
40 (обычно рибосомы не содержат двух или нескольких одинаковых белков).
Рибосомные белки характеризуются глобулярной компактной конформацией с развитой
вторичной и третичной структурой; они занимают преимущественно периферическое
положение в ядре, состоящем из рРНК. Плотность упаковки рРНК в рибосомах
достаточно высока.
По-видимому, рРНК определяет
основные структурные и функциональные свойства рибосом, в частности
обеспечивает целостность рибосомных субъединиц, обусловливает их форму и ряд
структурных особенностей. Специфическая пространственная структура рРНК
детерминирует локализацию всех рибосомных белков, играет ведущую роль в
организации функциональных центров рибосом.
3.
Принципы функционирования, роль РНК
Биосинтез белка имеет два
аспекта: химический и генетический. Принципиальным моментом является то, что в
природе белок строится из аминокислот не посредством их прямой конденсации с
освобождением воды или одновременной полимеризации на матрице, а путем
последовательного добавления аминокислотных остатков к одному из концов
растущей полипептидной цепи (удлинения) с одновременным сканированием
матричного полинуклеотида (мРНК), задающего порядок добавления различных
аминокислотных остатков. Три последовательные химические реакции приводят к
включению (добавлению) аминокислоты в полипептидную цепь строящегося белка.
Разделение
декодирующей и энзиматической функций между субчастицами
Трансляция начинается с того, что мРНК, синтезируемая на ДНК в качестве копии одной из двух цепей последней, связывается с рибосомной частицей. При этом рибосомная частица (у прокариот прямо и непосредственно, а у эукариот после некоторого скольжения вдоль некодирующей части мРНК) специфически взаимодействует с началом кодирующей нуклеотидной последовательности мРНК. Этап связывания мРНК с рибосомной частицей и нескольких последующих событий, приводящих к образованию первой пептидной связи, называется инициацией трансляции. Вслед за инициацией рибосома последовательно "читает" цепочку мРНК по тройкам (триплетам) нуклеотидов по направлению к 3"-концу, наращивая (удлиняя) полипептидную цепочку аминокислотными остатками; этот этап собственно трансляции называется элонгацией. Наконец, достигнув специального нуклеотидного триплета - стоп-кодона, или кодона терминации, - рибосома освобождает синтезированную полипептидную цепочку белка: происходит терминация трансляции.
Генетические функции малой рибосомной субчастицы.
Характерным моментом инициации трансляции является то, что на этом этапе
участвуют не целые рибосомы, а их отдельные субчастицы. Другими словами, для
того чтобы инициировать трансляцию, рибосома должна быть диссоциирована на две
составляющие ее неравные субчастицы. Для этого клетка располагает специальными
механизмами, обеспечивающими диссоциацию рибосом после терминации трансляции.
Именно малая субчастица рибосомы (30S у прокариот и 40S у эукариот), и только
она, связывается с мРНК, то есть служит первичным приемником генетической
информации для белоксинтезирующего аппарата. Лишь впоследствии, при завершении
этапа инициации трансляции, к ней присоединяется большая субчастица (50S у
прокариот и 60S у эукариот), образуя полную рибосомную частицу (70S у прокариот
и 80S у эукариот), которая и будет производить элонгацию.

В процессе элонгации рибосома удерживает мРНК и движется относительно ее
(или протягивает ее сквозь себя) в направлении от 5"-конца к 3"-концу.
Удержание мРНК на рибосоме есть целиком и полностью функция малой рибосомной
субчастицы, в то время как большая субчастица с мРНК никак не взаимодействует.
Соответственно последовательное сканирование кодирующей последовательности мРНК
(считывание генетической информации) в ходе элонгации осуществляется на малой
субчастице транслирующей рибосомы.
Механизм потриплетного сканирования мРНК в ходе элонгации предполагает участие
молекул тРНК, которые взаимодействуют прежде всего с малой рибосомной
субчастицей. Малая субчастица в составе полной транслирующей рибосомы имеет два
тРНК-связывающих участка, обозначаемых как аминоацил-тРНК-связывающий участок
(А-участок) и пептидил-тРНК-связывающий участок (Р-участок). На этапе элонгации
Р-участок всегда занят остатком тРНК.
Рассмотрение элементарного акта элонгации удобно начать с момента, когда Р-участок занят молекулой пептидил-тРНК (тРНК, несущая растущую полипептидную цепь), а А-участок вакантен и содержит лишь некий нуклеотидный триплет (кодон) мРНК, пока не взаимодействующий ни с каким триплетом (антикодоном) тРНК (рис. 1, состояние I). Такая рибосома готова (компетентна) связать аминоацил-тРНК, антикодон которой комплементарен триплету (кодону), установленному в А-участке. При наличии около рибосомы такой аминоацил-тРНК происходит первый шаг элементарного элонгационного цикла - кодонспецифическое связывание аминоацил-тРНК с А-участком. Теперь рибосома несет "старую" пептидил-тРНК в Р-участке и новоявленную аминоацил-тРНК в А-участке, которые расположены рядом, бок о бок (рис. 1, состояние II). Следовательно, в результате кодон-антикодонового взаимодействия мРНК с тРНК на малой субчастице рибосомы произошло декодирование триплета мРНК: именно тот аминокислотный остаток, который был привешен к тРНК с комплементарным антикодоном, оказался в рибосоме.
Далее молекулы пептидил-тРНК и аминоацил-тРНК, расположенные рядом в
рибосоме, реагируют друг с другом: пептидильный остаток переносится на
аминогруппу молекулы аминоацил-тРНК. Это второй шаг элементарного элонгационного
цикла - транспептидация, когда полипептидная цепь удлиняется на одну
аминокислоту - на ту, которую принесла тРНК, связавшаяся с А-участком. А сама
тРНК, принесшая эту аминокислоту, так и осталась с ней связанной и, таким
образом, связанной с удлиненным полипептидом (рис. 1, состояние III). В этом
состоянии, однако, новообразованная пептидил-тРНК - точнее, ее остаток тРНК -
занимает "не положенный ей" А-участок, а "сидит" в
Р-участке деацилированная (без пептидильного или аминоацильного остатков) тРНК.
Такое состояние называется претранслокационным. Дальше элонгация идти не может,
пока не осуществится третий шаг элонгационного цикла - транслокация, которая
выбросит деацилированную тРНК из Р-участка и переведет пептидил-тРНК из
А-участка в Р-участок вместе со связанным с ней кодоном мРНК. В результате в
освободившемся А-участке на малой рибосомной субчастице установится следующий
(новый) кодон мРНК.
Цикл завершился, приведя к
образованию одной пептидной связи и соответствующему удлинению растущего полипептида
на одну аминокислоту, с одной стороны, и к прочтению одного кодона мРНК и
перемещению мРНК на один триплет - с другой. Повторение таких элементарных
циклов и создает процесс элонгации.
Таким образом, малая рибосомная
субчастица в изолированном состоянии воспринимает копию гена в форме мРНК и
инициирует процесс ее трансляции, а в ходе трансляции малая субчастица полной
рибосомы удерживает мРНК на себе, декодирует ее с помощью тРНК и
последовательно перебирает ее кодоны и тРНК, используя механизм транслокации.
Так как все это операции с генетическим материалом, то указанные функции малой
рибосомной субчастицы могут быть определены как генетические.

Рис.
2: Реакция транспептидации, катализируемая пептидилтрансферазным центром
большой рибосомной субчастицы
Энзиматические функции большой рибосомной субчастицы.
Когда пептидил-тРНК занимает Р-участок, а аминоацил-тРНК - А-участок на малой субчастице рибосомы (см. рис. 1, состояние II) , концы остатков тРНК с присоединенными к ним аминоацильными остатками взаимодействуют с большой субчастицей рибосомы. Участок этого взаимодействия на большой субчастице является пептидилтрансферазным центром рибосомы: он катализирует реакцию транспептидации между пептидил-тРНК и аминоацил-тРНК, то есть перенос карбоксильной группы пептидильного остатка на аминогруппу аминоацил-тРНК (рис. 2). В результате образуется новая пептидная связь, и пептидильный остаток становится на одну аминокислоту длиннее. Таким образом, большая субчастица транслирующей рибосомы выступает здесь как фермент, ответственный за образование пептидных связей и в целом за синтез (элонгацию) полипептидной цепи. Это главная энзиматическая функция рибосомы.
Следует отметить, что никакого отдельного от рибосомы белка-фермента, катализирующего образование пептидных связей на рибосоме, не существует. Не найдено и никакого специального белка в составе рибосомы, который бы обладал такой энзиматической функцией. Транспептидация катализируется пептидилтрансферазным центром самой рибосомы как интегральной частью большой рибосомной субчастицы, и основной вклад в организацию центра вносит, по-видимому, рибосомная РНК субчастицы.
Кроме катализа реакции транспептидации большая рибосомная субчастица определенным образом участвует в энзиматическом расщеплении (гидролизе) гаунозинтрифосфата (ГТФ) в процессе трансляции. Дело в том, что, как видно на рис. 1, первый и третий шаги элонгационного цикла идут с участием специальных нерибосомных белков - так называемых факторов элонгации EF1 и EF2. Эти белки являются катализаторами соответствующих нековалентных переходов - связывания аминоацил-тРНК и транслокации. Для такого катализа необходимым оказывается сопряженный гидролиз ГТФ. Именно большая рибосомная субчастица взаимодействует с факторами элонгации и индуцирует гидролиз ГТФ на них. Хотя сам ГТФазный центр находится не на рибосомной субчастице, а на белке - факторе элогнации, ее временная ассоциация с фактором существенна для формирования активного энзиматического ГТФазного центра.
Таким образом, существует четкое разделение труда между двумя неравными
субчастицами рибосомы: малая субчастица выполняет генетические функции, будучи
ответственной за прием и декодирование генетической информации, в то время как
большая участвует в энзиматических реакциях в процессе трансляции.
Конформационная подвижность рибосомы
Работа рибосомы в качестве "лентопротяжного механизма"
последовательное прочитывание цепи мРНК от одного конца к другому) в ходе
элонгации и ее способность перебрасывать сравнительно большие молекулярные
массы (молекулы тРНК) из одного участка в другой в каждом элементарном
элонгационном цикле (см. рис. 1, шаг 3) предполагают ее механическую
подвижность. Взаимная подвижность двух рибосомных субчастиц может быть основным
видом крупноблочной подвижности рибосомы в ходе работы, и имеются
экспериментальные свидетельства в пользу такой подвижности. Кроме того,
существуют указания на подвижность "головки" малой рибосомной
субчастицы относительно ее "тела" и на подвижность палочкообразного
бокового выступа большой рибосомной субчастицы.
Рибосома при прохождении элонгационного цикла осциллирует между двумя
конформационными состояниями: закрытым (сомкнутым) и открытым (разомкнутым). В
сомкнутом состоянии рибосомные лиганды (тРНК) зажаты между субчастицами,
связаны максимальным количеством контактов с рибосомой и не имеют
внутририбосомной подвижности. В разомкнутом состоянии рибосомы лиганды более
подвижны, контакты с рибосомой менее полны, и имеется возможность их входа и
выхода из рибосомы. Так, на первом этапе связывания аминоацил-тРНК рибосома
должна быть открыта для приема лиганда. Возможно, это открытое состояние
фиксируется фактором элонгации EF1. Далее EF1 уходит, рибосомные субчастицы
плотно смыкаются, и аминоацильный конец связавшейся аминоацил-тРНК вступает в
контакт с пептидилтрансферазным центром большой субчастицы. В сомкнутом
состоянии пептидил-тРНК и аминоацил-тРНК тесно сближены, и между ними
происходит реакция транспептидации. Теперь, чтобы выбросить деацилированную
тРНК из рибосомы и дать свободу для перемещения остатка тРНК молекулы
пептидил-тРНК из А-участка в Р-участок, рибосому надо приоткрыть, в частности
путем раздвигания субчастиц. Это может осуществляться фактором
элонгации EF2. После ухода EF2 с рибосомы она снова смыкается и ждет прихода
очередной аминоацил-тРНК с фактором элонгации EF1.
Процесс периодического
смыкания-размыкания рибосомы является энергозависимым: факторы элонгации EF1 и
EF2 взаимодействуют с рибосомой только будучи связанными с ГТФ (согласно
модели, при этом взаимодействии происходит открывание рибосомы), а
взаимодействие с рибосомой наводит ГТФазную активность, ГТФ гидролизуется,
фактор элогации теряет сродство к рибосоме и уходит, и рибосома закрывается.
Таким образом, на каждое смыкание-размыкание рибосомы расходуется одна молекула
ГТФ. Так как в каждом элонгационном цикле рибосома смыкается-размыкается
дважды, то две молекулы ГТФ расходуются на каждый цикл. Это есть энергетическая
плата за эффективное (быстрое и надежное) функционирование рибосомы как
молекулярной машины.
4.
Список литературы
1. Спирин А.С. Принципы структуры рибосом // Соросовский Образовательный Журнал. 1998. N 11. С. 65-70.
2. Спирин А.С. Молекулярная биология: Структура рибосомы и биосинтез белка. М.: Высш. шк., 1986. 300 с.
3. Спирин А.С. О механизме работы рибосомы: Гипотеза смыкания-размыкания субчастиц // Докл. АН СССР. 1968. Т. 179. С. 1467-1470.
Строение рибосомы. Рибосомы обнаружены в клетках всех организмов. Это микроскопические тельца округлой формы диаметром 15-20 нм. Каждая рибосома состоит из двух неодинаковых по размерам частиц, малой и большой.
В одной клетке содержится много тысяч рибосом, они располагаются либо на мембранах гранулярной эндоплазматической сети, либо свободно лежат в цитоплазме. В состав рибосом входят белки и РНК. Функция рибосом - это синтез белка. Синтез белка - сложный процесс, который осуществляется не одной рибосомой, а целой группой, включающей до нескольких десятков объединенных рибосом. Такую группу рибосом называют полисомой. Синтезированные белки сначала накапливаются в каналах и полостях эндоплазматической сети, а затем транспортируются к органоидам и участкам клетки, где они потребляютя. Эндоплазматическая сеть и рибосомы, расположенные на ее мембранах, представляют собой единый аппарат биосинтеза и транспортировки белков.
Химический состав рибосом В составе рибосом эукариотического типа 4 молекулы рРНК и около 100 молекул белка, прокариотического типа - 3 молекулы рРНК и около 55 молекул белка. Во время биосинтеза белка рибосомы могут «работать» поодиночке или объединяться в комплексы - полирибосомы (полисомы). В таких комплексах они связаны друг с другом одной молекулой иРНК. Прокариотические клетки имеют рибосомы только 70S-типа. Эукариотические клетки имеют рибосомы как 80S-типа (шероховатые мембраны ЭПС, цитоплазма), так и 70S-типа (митохондрии, хлоропласты).Субъединицы рибосомы эукариот образуются в ядрышке. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка.
Функция рибосом: сборка полипептидной цепочки (синтез белка).
Свободные рибосомы, полирибосомы, их связь с другими структурными компонентами клетки.
Различают единичные рибосомы и комплексные рибосомы (полисомы). Рибосомы могут располагаться свободно в гиалоплазме и быть связанными с мембранами эндоплазматической сети. Свободные рибосомы образуют белки в основном на собственные нужды клетки, связанные обеспечивают синтез белков “” на экспорт”.
Конец работы -
Эта тема принадлежит разделу:
Гистология
Гистология от греч histos ткань logos учение наука о строении развитии и жизнедеятельности тканей живых организмов.. Становление гистологии тесно связано с развитием микроскопической техники и.. В истории учения о тканях и микроскопическом строении органов выделяют два периода домикроскопический и..
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Все темы данного раздела:
Уровни организации живой материи в целостном организме. Их морфофункциональные особенности и коррелятивные связи
1. Молекулярный. Любая живая система проявляется на уровне взаимодействия биологических макромолекул: нуклеиновых кислот, полисахаридов, а также других важных органических веществ.
2. Клет
Методы исследования
В современной гистологии, цитологии и эмбриологии применяются разнообразные методы исследования, позволяющие всесторонне изучать процессы развития, строения и функции клеток, тканей и органов.
Органеллы цитоплазмы клетки. Определение, их функции. Мембранные и немембранные органеллы. Внутренний сетчатый аппарат, структура и функция
Органеллы
Органеллы – постоянные структурные элементы цитоплазмы клетки, имеющие специфическое строение и выполняющие определенные функции.
Классификация органелл:
1) общ
Включения(все про них, характеристики)
Включения - непостоянные структурные компоненты цитоплазмы.
Классификация включений:
трофические:
лецитин в яйцеклетках;
гликоген;
липиды, имеются почти
Ядро(все про него)
Ядро-компонент клетки,который содержит генетт.материал.
Функции ядра: хранение,реализация, передача генет.информации
Ядро состоит из:
Кариолеммы-ядерной оболочки
Способы репродукции клеток. Митоз, его смысл биологический. Эндорепродукция
Различают два основных способа размножения клеток: митоз (кариокенез) – непрямое деление клеток, которое присуще в основном соматическим клеткам;Биологический смысл митоза-из одной диплоидной м
Жизненный цикл клетки, его этапы
Положения клеточной теории Шлейдена-Шванна
Все животные и растения состоят из клеток.
Растут и развиваются растения и животные путём возникновения новых клет
1. Ткань – исторически (филогенетически) сложившаяся система клеток и неклеточных структур, обладающая общностью строения, а иногда и происхождения, и специализированная на выполнение определенных
Покровный эпителий
Покровные эпителии
В соответствии с морфологической классификацией различают несколько основных типов покровного эпителия,как многослойного, так и однослойного. При этом для многослойных э
Эритроциты
Эритроциты у человека и млекопитающих представляют собой безъядерные клетки, утратившие в процессе фило- и онтогенеза ядро и большинство органелл. Эритроциты являются высокодифференцированными пост
Кровь,как ее ткань,ее форменные элементы.Кровяные пластинки(тромбоциты),их количество.размеры.строение.функции.продолжительность жизни
Кровь - жидкая соединительная ткань, циркулирующая в кровеносной системе тела животного. У всех позвоночных кровь имеет красный цвет (от ярко- до тёмно-красного), которым она обязана гемоглобину, с
Мышца как орган. Микроскопическое строение мыщц. Мион. Связь мышц сухожилием
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его часте
Сердечная мыш. ткань (поперечнополосатая мышечная ткань целомического типа) встречается в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов. Её кл-ки (сердечные миоцит
Мозжечок. Строениеи функциональная характеристика. Нейронный состав коры мозжечка и глиоцыты. Межнейронные связи
Мозжечок. Представляет собой центральный орган равновесия и координации движений. Он связан со стволом мозга афферентными и эфферентными проводящими пучками, образующими в совокупности три пары нож
Капилляры. Строение. Органоспецифичность капилляров. Понятие о гистогематическом барьере. Венулы, их функциональное значение и строение
Микроциркуляторное русло - система мелких сосудов, включающая артериолы, гемокапилляры, венулы, а также артериоловенулярные анастомозы. Этот функциональный комплекс кровеносных сосудов, окруженный
Вены. Особенности строения вен различного типа. Органные особенности вен
Вены - осуществляют отток крови от органов, участвуют в обменной и депонирующей функциях. Различают поверхностные и глубокие вены. Вены широко анастомозируют, образуя в органах сплетения.
Эмбриогенез органа зрения
Глазное яблоко формируется из нескольких источников. Сетчатка является производным нейроэктодермы и представляет собой парное выпячивание стенки diencephalon в виде однослойного пузырька на ножке.
Вкусовая сенсорная система. Орган вкуса
Орган вкуса (organum gustus) - периферическая часть вкусового анализатора представлен рецепторными эпителиальными клетками во вкусовых почках (caliculi gustatoriae). Они воспринимают вкусовые раздр
Эмбриогенез органа слуха
Внутреннее ухо. Первым из структур внутреннего уха развивается перепончатый лабиринт. Исходным материалом для него является эктодерма, лежащая на уровне заднего мозгового пузыря. Впячиваясь в подле
Эндокринная система
Гуморальная регуляция, гормоны, классификация эндокринных желез
При изучении эпителиальных тканей организма в классификации, наряду с покровным эпителием, выделялся железистый эпителий, в
Гипоталамус
Гипоталамус - высший нервный центр регуляции эндокринных функций. Этот участок промежуточного мозга является также центром симпатического и парасимпатического отделов вегетативной нервной системы.
Половые гормоны
Половые гормоны - это гормоны, вырабатываемые мужскими и женскими половыми железами и корой надпочечников. Все половые гормоны по химическому строению являются стероидами. К половым гормонам от
Развитие щитовидной железы
Зачаток щитовидной железы появляется на 4-й неделе эмбриогенеза в виде выпячивания вентральной стенки глоточной кишки между 1-й и 2-й парами жаберных карманов. Выпячивание это превращается в эпител
Околощитовидные железы
Источники развития.
Околощитовидные железы - производные 3-й и IV-й пар жаберных карманов, эпителиальная выстилка которых имеет прехордальный генез. На 5-6-й неделе эмбриогенеза образуются
Надпочечники
Надпочечники представляют собой парные железы, состоящие из коркового и мозгового вещества. Каждая из этих частей является самостоятельной железой внутренней секреции, вырабатывающей свои гормоны -
Эпифиз
Эпифиз (верхний мозговой придаток, пинеальная, или шишковидная, железа) расположен между передними буграми четверохолмия. Это нейроэндокринный орган, регулирующий физиологические ритмы, так как сек
А. Ротовая полость
Слизистая оболочка ротовой полости состоит из многослойного плоского эпителия кожного типа, развивающегося из прехордальной пластинки, и собственной соединительнотканной пластинки. Степень развития
Большие слюнные железы
Помимо множества мелких слюнных желез, расположенных в слизистой оболочке щек, и желез языка, в полости рта находятся большие слюнные железы (околоушные, поднижнечелюстные и подъязычные), являющиес
Пищевод
Источником развития эпителия пищевода является материал прехордальной пластинки. Остальные ткани стенки пищевода за некоторым исключением развиваются из мезенхимы. Выстилка пищевода сначала предста
Желудок
Средний, или гастроэнтеральный, отдел пищеварительной трубки включает желудок, тонкую и толстую кишки, печень и желчный пузырь, поджелудочную железу. В этом отделе происходят переваривание пищи под
Тонкая кишка
В тонкой кишке различают три переходящих друг в друга отдела: двенадцатиперстную, тощую и подвздошную кишки. В тонкой кишке происходят дальнейшее переваривание пищи, предварительно обработанной в р
Толстая кишка
В толстой кишке происходят интенсивное всасывание воды, переваривание клетчатки с участием бактериальной флоры, выработка витамина К и комплекса витаминов В, выделение ряда веществ, например, солей
Железы пищеварительной системы. Поджелудочная железа
Поджелудочная железа состоит из экзокринной и эндокринной частей. Экзокринная часть выполняет внешнесекреторную функцию, связанную с выработкой панкреатического сока. Он содержит пищеварительные фе
Печень. Желчный пузырь
Печень - самая крупная железа человека - ее масса составляет около 1,5 кг. Она выполняет многообразные функции и является жизненно важным органом. Чрезвычайно важными для поддержания жизнеспособнос
Гемопоэз
Дифференциация - это стойкое структурно-функциональное преобразование клеток в различные специализированные клетки. Дифференцировка клеток биохимически связана с синтезом специфических белков, а ци
Красный костный мозг
Красный костный мозг
Красный костный мозг - центральный гемопоэтический орган. В нем находится основная часть стволовых кроветворных клеток и происходит развитие клеток миелоидного и лимфо
Тимус. Развитие тимуса. Строение тимуса
Тимус - центральный орган лимфоидного кроветворения и иммунной защиты организма. В тимусе происходит антигеннезависимая дифференцировка костномозговых предшественников Т-лимфоцитов в иммунокомпетен
Селезенка
СТРОМА
плотная строма: капсула и септы (септы в селезенке называются трабекулами) образованы плотной волокнистой соединительной тканью, где имеется много эластических волокон, встречаются
Лимфатические узлы
СТРОМА
плотная строма: капсула и септы образованы РВСТ
мягкая строма: ретикулярная ткань; в корковом веществе - в лимфоидных фолликулах имеется особая разновидность клеток ретикул
типа - плоские, или респираторные
Покрывают бо́льшую часть (95-97 %) поверхности альвеол, являются компонентом аэрогематического барьера, через них осуществляется газообмен. Имеют неправильную форму и истончённую цитоплазму (м
Сурфактантная система легких
Вверху справа - кровеносный капилляр, содержащий эритроцит. Назальная мембрана капилляра слилась с мембраной лежащего вверху плоского эпителия, образуя в отмеченных участках. Сурфактантная система
Железы кожи
Потовые железы участвуют в терморегуляции, а также в экскреции продуктов обмена, солей, лекарственных веществ, тяжелых металлов (усиливается при почечной недостаточности).
Потовые
Особенности кровоснабжения почек
Каждая почка имеет достаточно своеобразную сосудистую сеть. В ворота Почки входит так называемая почечная артерия (a. renalis). Почечная артерия разветвляется на несколько так называемых сегментарн
Мочеточники - парный орган мочевой системы человека
Характеристики
Правый и левый мочеточники
Представляют собой протоки длиной от 27 до 30 см, диаметром от 5 до 7 мм
Прощупать через живот невозможно
Внешняя стенк
Яичники
Анатомически яичник представлен в виде овоидного тела длиной 2,5 – 5,5см, шириной 1,5 -3,0см. Масса обоих яичников у новорожденных в среднем составляет 0,33г, у взрослых – 10,7г.
Функция:
Яичник взрослой женщины
С поверхности орган окружен белочной оболочкой (tunica albuginea), образованнойплотной волокнистой соединительной тканью, покрытой мезотелием брюшины. Свободная поверхность мезотелия снабжен
Менструальная фаза
В этой фазе происходит отторжение (десквамация) функционального слоя эндометрия матки, которое сопровождается кровотечением.
В момент окончания менструации эндометрий представлен то
Органеллы клетки, состоящие из белков и РНК и отвечающие за синтез белков, называются рибосомами. Количество рибосом в одной клетке сильно варьирует в зависимости от потребностей и может достигать нескольких миллионов.
Строение
Важнейшей органеллой клетки является ядро. Оно содержит генетическую информацию и ядрышко, где образуются рибосомы. Синтезированные рибосомы через поры ядерной мембраны попадают либо на эндоплазматическую сеть, либо в цитоплазму. В зависимости от расположения в эукариотической клетке выделяют два вида рибосом:
- связанные - располагаются на эндоплазматической сети (шероховатый вид);
- свободные - располагаются в цитозоле.
Гладкая ЭПС образуется после освобождения от рибосом. В растительных клетках гладкая ЭПС формирует провакуоли, из которых затем образуются вакуоли.

Рис. 1. Расположение рибосом в клетке.
Рибосомы - немембранные органеллы, имеющие округлую форму и состоящие из двух частей - субъединиц (большой и малой), каждая из которых представляет собой смесь рибосомальной РНК (рРНК) и белков. С химической точки зрения рибосома - нуклеопротеид, состоящий из нуклеиновых кислот и протеинов.

Рис. 2. Строение рибосом.
Связанные и свободные рибосомы называются цитоплазматическими рибосомами. Также существуют собственные рибосомы митохондрий и пластид. Они отличаются меньшим количеством белков и рРНК.
Различают четыре разновидности молекул РНК рибосомы:
ТОП-3 статьи которые читают вместе с этой
- 18S-РНК - содержит 1900 нуклеотидов;
- 5S-РНК - содержит 120 нуклеотидов;
- 5,8S-РНК - состоит из 160 нуклеотидов;
- 28S-РНК - состоит из 4800 нуклеотидов.
Малая частица рибосомы образована 30-35 белками и 18S-РНК. В большую субчастицу входит 45-50 белков и 5S-, 5,8S-, 28S-РНК.
В нерабочем состоянии части рибосом разъединены. Они соединяются с помощью информационной (матричной) РНК, обхватывая её с двух сторон. При синтезе белка рибосомы объединяются, образуя комплексы - полисомы или полирибосомы, связанные мРНК и напоминающие бусины на нитке.
Рибосомы прокариот меньше, чем эукариот. Диаметр рибосом клетки человека, животных, растений и грибов - 25-30 нм, бактерий - 15-20 нм.
Синтез белка
Главная функция рРНК - синтез белка и аминокислот.
Биосинтез белков включает два процесса:
- транскрипцию;
- трансляцию.
Транскрипция происходит с участием ДНК. Генетическую информацию считывает фермент РНК-полимераза, образуя мРНК. Далее начинается процесс трансляции, происходящий на рибосомах.
Этот процесс разделяется на три этапа:
- инициацию - начало синтеза;
- элонгацию - биосинтез;
- терминацию - завершение синтеза, отделение рибосомы.
При инициации происходит сборка рибосомы. Контактные части субъединиц называются активными центрами, между которыми располагается :
- мРНК в качестве «шаблона» синтеза;
- тРНК, осуществляющая перенос аминокислот на синтезируемую цепь;
- синтезируемый пептид, состоящий из аминокислот.
В процессе элонгации происходит удлинение полипептидной цепи за счёт присоединения аминокислот. Цепь отсоединяется от рибосомы на стадии терминации благодаря стоп-кодону - единицы генетического кода, шифрующего прекращение синтеза белка.

Рис. 3. Общая схема синтеза белка на рибосоме.
Биосинтез требует энергетических затрат. При присоединении одной аминокислоты расходуется по две молекулы АТФ (аденозинтрифосфата) и ГТФ (гуанозинтрифосфата). Кроме того, ГТФ тратится на процессы инициации и терминации.
Что мы узнали?
Из урока 9 класса кратко узнали о строении и функции рибосомы. Это важные органоиды клетки, осуществляющие биосинтез белка путём считывания информации с мРНК. Рибосомы образованы двумя частями (большой и малой), каждая из которых состоит из рибонуклеиновой кислоты и белков.
Тест по теме
Оценка доклада
Средняя оценка: 4.6 . Всего получено оценок: 116.
Рибосома(от «РНК» и soma – тело) – клеточный немембранный органоид, осуществляющий трансляцию (считывание кода мРНК и синтез полипептидов).
Рибосомы эукариот расположены на мембранах эндоплазматической сети (гранулярная ЭС) и в цитоплазме. Прикрепленные к мембранам рибосомы синтезируют белок «на экспорт», а свободные рибосомы – для нужд самой клетки. Различают 2 основных типа рибосом – прокариотные и эукариотные. В митохондриях и хлоропластах также имеются рибосомы, которые близки к рибосомам прокариот.
Рибосома состоит из двух субъединиц – большой и малой. У прокариотических клеток они обозначены 50S и 30S субъединицы, у эукариотических – 60S и 40S. (S – коэффициент, который характеризует скорость осаждения субъединицы при ультрацентрифугировании). Субъединицы эукариотических рибосом образуются путем самосборки в ядрышке и через поры ядра поступают в цитоплазму.
Рибосомы в клетках эукариот состоят из четырех нитей РНК (три молекулы рРНК в большой субъединице и одна молекула рРНК – в малой) и примерно 80 разных белков, т.е представляют собой сложнейший комплекс из молекул, скрепленных слабыми, нековалентными связями. (Рибосомы в клетках прокариот состоят из трех нитей РНК; две нити рРНК находятся в большой субъединице и одна рРНК – в малой). Процесс трансляции (биосинтеза белка) начинается со сборки активной рибосомы. Этот процесс называется инициацией трансляции. Сборка происходит строго упорядоченным образом, что обеспечивается функциональными центрами рибосом. Все центры находятся на контактирующих поверхностях обеих субъединиц рибосомы. Каждая рибосома работает как большая биохимическая машина, а точнее, как суперфермент, который, во-первых, правильно ориентирует участников (мРНК и тРНК) процесса друг относительно друга, а во-вторых, катализирует реакции между аминокислотами.
Активные центры рибосом:
1)центр связывания мРНК (М-центр);
2) пептидильный центр (П-центр). С этим центром в начале процесса трансляции связывается инициирующая тРНК; на последующих стадиях трансляции из А-центра в П-центр перемещается тРНК, удерживающая синтезированную часть пептидной цепи;
3)аминокислотный центр (А-центр) – место связывания кодона мРНК с антикодоном тРНК, несущей очередную аминокислоту.
4)пептидилтрансферазный центр (ПТФ-центр): он катализирует реакцию связывания аминокислот. При этом образуется еще одна пептидная связь, и растущий пептид удлиняется на одну аминокислоту.

Схема синтеза белка на рибосомах гранулярной эндоплазматической сети.
(рис. из книги биология клетки, том II )

Схематическое изображение полирибосомы. Синтез белка начинается со связывания малой субчастицы, в месте расположения AUG -кодона в молекуле информационной (матричной РНК) (рис. из книги биология клетки, том II ).
Эндоплазматическая сеть
Эндоплазматическая сеть (син. эндоплазматический ретикулум) – органоид эукариотической клетки. В клетках разного типа и при различных функциональных состояниях этот компонент клетки может выглядеть по-разному, но во всех случаях – это лабиринтная протяженная замкнутая мембранная структура, построенная из сообщающихся трубкообразных полостей и мешочков, называемых цистернами. Снаружи от мембран эндоплазматической сети находится цитозоль (гиалоплазма, основное вещество цитоплазмы), а просвет эндоплазматической сети представляет собой замкнутое пространство (компартмент), сообщающееся посредством везикул (транспортных пузырьков) с комплексом Гольджи и внешней для клетки средой. Эндоплазматическая сеть делится на две функционально различные структуры: гранулярную (шероховатую) эндоплазматическую сеть и гладкую(агранулярную) эндоплазматическую сеть.
Гранулярная эндоплазматическая сеть, в клетках секретирующих белок, представлена системой многочисленных плоских мембранных цистерн с рибосомами на наружной поверхности. Комплекс мембран гранулярной эндоплазматической сети связан с наружной мембраной оболочки ядра и перинуклеарной (околоядерной) цистерной.
В гранулярной эндоплазматической сети происходит синтез белков и липидов для всех мембран клетки, синтезируются ферменты лизосом, а также осуществляется синтез секретируемых белков, т.е. предназначенных для экзоцитоза. (Остальные белки синтезируются в цитоплазме на рибосомах, не связанных с мембранами ЭС.) В просвете гранулярной ЭС белок окружается мембраной, и образующиеся пузырьки отделяются (отпочковываются) от несодержащих рибосомы областей ЭС, которые и доставляют содержимое в другую органеллу – комплекс Гольджи – путем слияния с ее мембраной.
Та часть ЭС, на мембранах которой рибосомы отсутствуют, называется гладким эндоплазматическим ретикулумом. Гладкая эндоплазматическая сеть не содержит уплощенных цистерн, а представляет собой систему анастомозирующих мембранных канал
ов, пузырьков и трубочек. Гладкая сеть является продолжением гранулярной, однако не содержит рибофоринов – гликопротеиновых рецепторов, с которыми соединяется большая субъединица рибосом и поэтому не связана с рибосомами.
Функции гладкой эндоплазматической сети многообразны и зависят от типа клеток. Гладкая эндоплазматическая сеть участвует в метаболизме стероидных, например, половых гормонов. В ее мембранах локализованы управляемые кальциевые каналы и энергозависимые кальциевые насосы. Цистерны гладкой эндоплазматической сети специализированы для накопления в них Са 2+ путем постоянного откачивания Са 2+ из цитозоля. Подобные депо Са 2+ существуют в скелетной и сердечной мышцах, нейронах, яйцеклетке, эндокринных клетках и др. Различные сигналы (например, гормоны, нейромедиаторы, факторы роста) влияют на активность клеток путем изменения концентрации внутриклеточного посредника – Са 2+ . В гладкой эндоплазматической сети клеток печени происходит обезвреживание вредных веществ, (например ацетальдегида, образующегося из алкоголя), метаболическая трансформация лекарств, образование большей части липидов клетки и их накопление, например при жировой дистрофии. В полости ЭС содержится много различных молекул-компонентов. Среди них имеют большое значение белки шапероны.

Шапероны (англ. букв. – пожилая дама, сопровождающая молодую девушку на балах) – семейство специализированных внутриклеточных белков, обеспечивающих быстрое и правильное сворачивание (фолдинг) вновь синтезированных молекул белка. Связывание с шаперонами препятствует агрегации с другими белками и тем самым создает условия для формирования вторичной и третичной структуры растущего пептида. Шапероны принадлежат к трем белковым семействам, так называемым белкам теплового шока (hsp 60, hsp 70, hsp 90). Синтез этих белков активируется при многих стрессах, в частности, при тепловом шоке (отсюда и название h eart shook protein – белок теплового шока, а цифра обозначает его молекулярную массу в килодальтонах). Эти шапероны предотвращают денатурацию белков при высокой температуре и др. экстремальных факторах. Связываясь с аномальными белками, восстанавливают их нормальную конформацию и тем самым повышают выживаемость организма при резком ухудшении физико-химических параметров среды.